

ヒトの眼の進化のターニングポイントを発見
”ホヤの光センサー”が埋める進化のmissing link
by Keiichi Kojima概要
ヒトを含む脊椎動物はカメラに似た眼をもっており、様々な光環境でものの形や色を見ることができます。眼には、外からの光を受容して神経の電気応答に変換する視細胞が含まれ、視細胞では光を受容するための光センサー(光受容タンパク質)が機能しています。視細胞の電気応答は脳へと伝えられ、最終的に”見えた”と実感できます。我々はこれまで、ヒトなど脊椎動物の視細胞に含まれる光受容タンパク質は、脊椎動物の先祖型のものに比べて、光を受けた後の光情報の増幅効率(シグナル増幅効率)が非常に高く、この性質は脊椎動物の光受容タンパク質が分子進化の過程で、新たに特別なアミノ酸残基を獲得したからであることを発見していました。しかし、このアミノ酸残基がどのような過程で先祖型の光受容タンパク質の中で獲得され機能するようになったかは明らかでありませんでした。 今回我々は、脊椎動物と最後に分かれた無脊椎動物であるホヤの光受容タンパク質を解析したところ(図1)、驚いたことに、この特別なアミノ酸残基は既に獲得されているが、従来から機能していたアミノ酸残基も同時に機能していることを発見しました。そこでホヤの光受容タンパク質の変異体解析により、新たに獲得されたアミノ酸残基をなくすと、無脊椎動物の光受容タンパク質と同様の光反応を示し、逆に従来機能していたアミノ酸残基をなくすと、脊椎動物の光受容タンパク質と同様の光反応を示すようになりました。しかし、変異体によって光シグナルの増幅効率は脊椎動物の光受容タンパク質のレベルまでは大きくならず、タンパク質構造のさらなる変化が必要であることもわかりました。 つまり、ホヤの光受容タンパク質は、進化の過程でシグナル増幅効率を上げるために必要な新規のアミノ酸残基を獲得しているが、まだ、完全な意味で脊椎動物の光受容タンパク質のようにはなっていない、ということがわかりました(図2)。まさに、脊椎動物の祖先からヒトの眼の高度な機能が創造される過程のmissing linkを埋める役割を果たしてくれました。ホヤは、我々のように発達した眼は持っていませんが、分子のレベルでは高度な視覚機能を進化させるための準備を既に始めていた、と言うことができます。 本研究成果は、2017年5月22日に米国科学アカデミー紀要オンライン版にて発表されました。

図1 カタユウレイボヤの幼生の眼点
ホヤの幼生は、明暗に応じて遊泳行動を変化させる。眼点の光受容タンパク質が、その行動変化に関わることが分かっている。
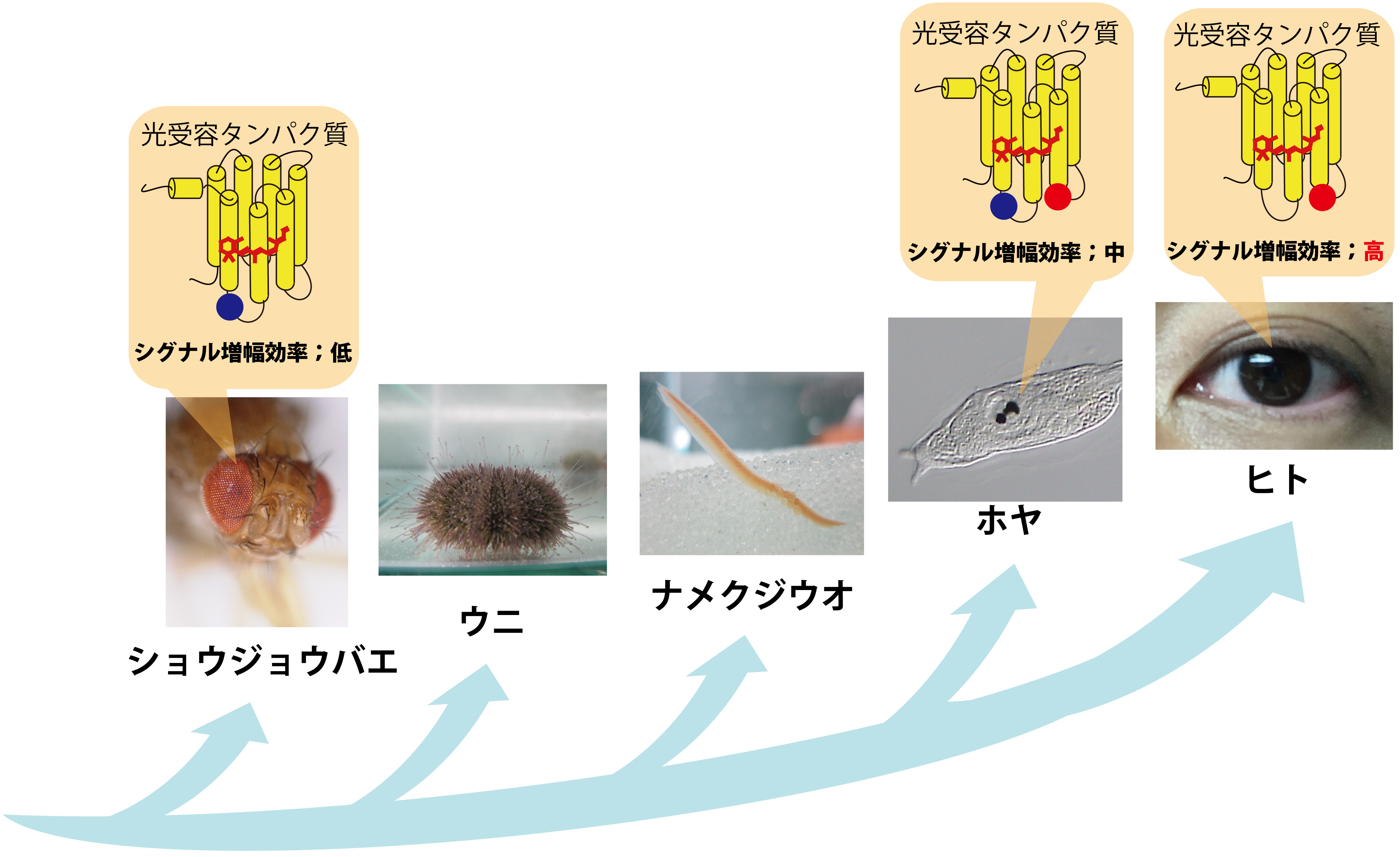
図2 脊椎動物の高度な視覚機能を支える光センサーの進化
視覚を担う光受容タンパク質のシグナル増幅効率の上昇が、脊椎動物の眼が劇的に進化することの一因になったと考えられる。赤丸は脊椎動物の光受容タンパク質の高いシグナル増幅効率に必要なアミノ酸残基を示し、青丸は従来機能していたアミノ酸残基を示している。
研究の背景
動物にとって、眼から得られる視覚の情報は外界の変化をとらえる上で非常に重要です。特にヒトでは、周りから得られる感覚情報の多くが視覚を通じて得られると言われ、脳の約半分が視覚の情報処理に関わっています。しかし、身の回りの動物を見ても、様々な形態の眼を持つものがいます。ヒトなど脊椎動物が持つカメラ眼とハエなど昆虫類が持つ複眼では、見た目が大きく異なっています。かつて、チャールズ・ダーウィンも眼の進化に大いに興味を持っていたことが、その著書『種の起源』の記述からも伺えます。そして、最近では、眼の形態的違いだけでなく、眼の中で機能する分子(タンパク質)に着目した研究も進んできました。 ヒトなど脊椎動物は高度に発達した眼を持ち、明るい場所でも暗い場所でも周りの環境を捉えることが可能です。この視覚機能を支えているのは、ロドプシン*1に代表される、光センサー(光受容タンパク質)であり、眼の網膜の視細胞で機能しています。この光受容タンパク質が光を受け、光シグナルが視細胞の電気応答に変換されます。そしてこの電気応答が最終的に脳へと伝達され、”見えた”と実感できます。 我々はこれまで、ヒトなど脊椎動物の眼が光への高い感受性を示すためにそこで機能する光受容タンパク質がどのように進化してきたのか、を解析してきました。カンブリア爆発の時期に他の動物群と分岐した脊椎動物の祖先は、他の動物と同様に、明暗を感じる程度の光受容器官しか持っていなかったと考えられます。そのため、そこで機能する光受容タンパク質も現在の脊椎動物のものと比べてシグナル増幅効率の悪いものであった、と推定できます。我々はこれまでの研究で、脊椎動物の祖先から進化する過程で、光受容タンパク質が新たに特別なアミノ酸残基を獲得することによってシグナル増幅効率を高めた、ことを発見していました。このような分子レベルの変化が、脊椎動物における眼の劇的な進化に寄与していると考えられますが、先祖型の光受容タンパク質の中で特別なアミノ酸残基がどのようにして獲得され機能するようになったかは明らかでありませんでした。
研究成果
本研究で我々は、脊椎動物に最も近縁な無脊椎動物であるホヤ(カタユウレイボヤ)に注目をしました。カタユウレイボヤは、成体では海底に付着していますが、幼生の段階では海中を遊泳します。そして、光環境の変化で幼生の動きが大きく変化します。幼生は、光受容細胞を含む眼点を持ち、そこで機能する光受容タンパク質が明暗による行動の変化に関わることが分かっていました(図1)。 このホヤの光受容タンパク質を調べたところ、脊椎動物の光受容タンパク質が高いシグナル増幅効率を示すために重要なアミノ酸残基を既に獲得していることを見いだしました。また、驚いたことに、無脊椎動物の光受容タンパク質で従来機能しているアミノ酸残基も同時に機能していることがわかりました。そこで、進化の道筋を戻すような形で、ホヤの光受容タンパク質において新たに獲得したアミノ酸残基を人為的になくした変異体を作製したところ、無脊椎動物の光受容タンパク質と同様の光反応を示しました。また、逆に進化の道筋を進ませるような形で、従来機能しているアミノ酸残基をなくしたところ、脊椎動物の光受容タンパク質と同様の光反応を示しました。しかし、この変異体でもシグナル増幅効率は脊椎動物の光受容タンパク質のレベルまでは大きくならず、タンパク質構造のさらなる変化が必要であることもわかりました。つまり、ホヤの光受容タンパク質は、進化の過程でシグナル増幅効率を上げるために必要な新規のアミノ酸残基を獲得しているが、まだ、完全な意味で脊椎動物の光受容タンパク質のようにはなっていない、ということがわかりました。まさに、脊椎動物の祖先からヒトの眼の高度な機能が創造される過程のmissing linkを埋める役割を果たしてくれました(図2)。ホヤは、我々のように発達した眼は持っていませんが、分子のレベルでは高度な視覚機能を進化させるための準備を既に始めていた、と言うことができます。
用語説明
*1ロドプシン:動物が持つ代表的な光センサーであり、タンパク質内部に発色団としてビタミンAの誘導体(レチナール)を結合する。網膜の視細胞において最初に光を受容し、視細胞内部へシグナルを伝えることにより、光シグナルを視細胞の電気応答に変換する。