
魚類の多様な光受容システムを導いたユニークな遺伝子重複
by Chihiro Fujiyabu概要
動物は光環境から様々な情報を得ています。視覚によりものの形や色を認識するだけではなく、時刻や季節の認識も行っています。これらの多様な光受容機能に広く関わる光センサータンパク質がオプシン類です。近年のゲノム解析の進展により、非常に多種多様なオプシン遺伝子が同定されています。その中で最も代表的なオプシンが眼の網膜で機能する視覚の光センサーであるロドプシンです。多くの脊椎動物においてロドプシン遺伝子は1種類であり、その翻訳領域にはイントロンが含まれます。しかし例外的に、真骨魚類という我々が普段目にする大多数の身近な魚が含まれる分類群は複数のロドプシン遺伝子を持つということが知られていました。真骨魚類の網膜で機能するロドプシン遺伝子はイントロンを含まず、網膜ではなく脳の松果体で機能するロドプシン遺伝子はイントロンを含みます。このことは、真骨魚類の祖先において、イントロンを含むロドプシン遺伝子からretroduplicationというユニークな遺伝子重複機構によってイントロンを含まないロドプシン遺伝子が生じ、前者は松果体に、後者は網膜に、それぞれで発現するように変化したという可能性を示しています。そこで我々は、真骨魚類の持つエキソン/イントロン構造の異なる2種類のロドプシンが獲得・分化した経緯を明らかにするため、条鰭綱において真骨魚類よりも古くに分岐したと考えられる古代魚(ポリプテルス・チョウザメ・ガー)のロドプシンを調べました。その結果から、条鰭綱の進化の初期、約4億年前に起こったretroduplicationによってイントロンなしロドプシン遺伝子が生じたのち、段階的な機能分化を経て2種類のロドプシンが眼と脳で使い分けられるようになったという進化モデルを提唱しました。本研究成果は2019年7月23日にScientific Reportsにて発表されました。
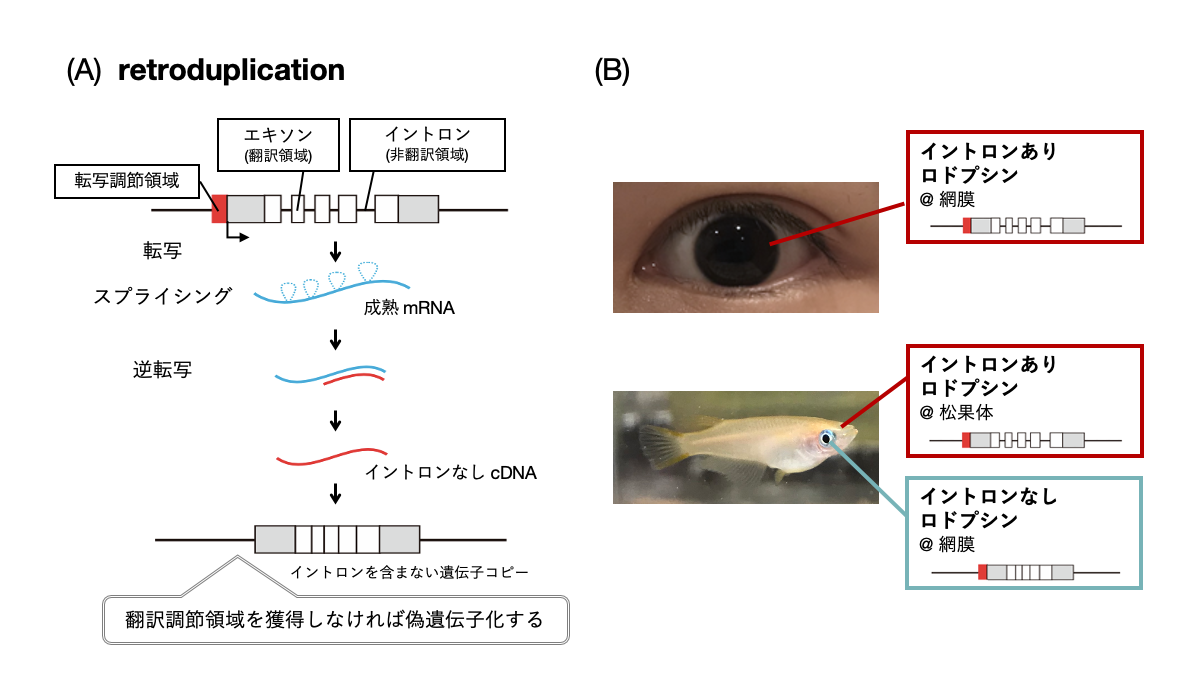
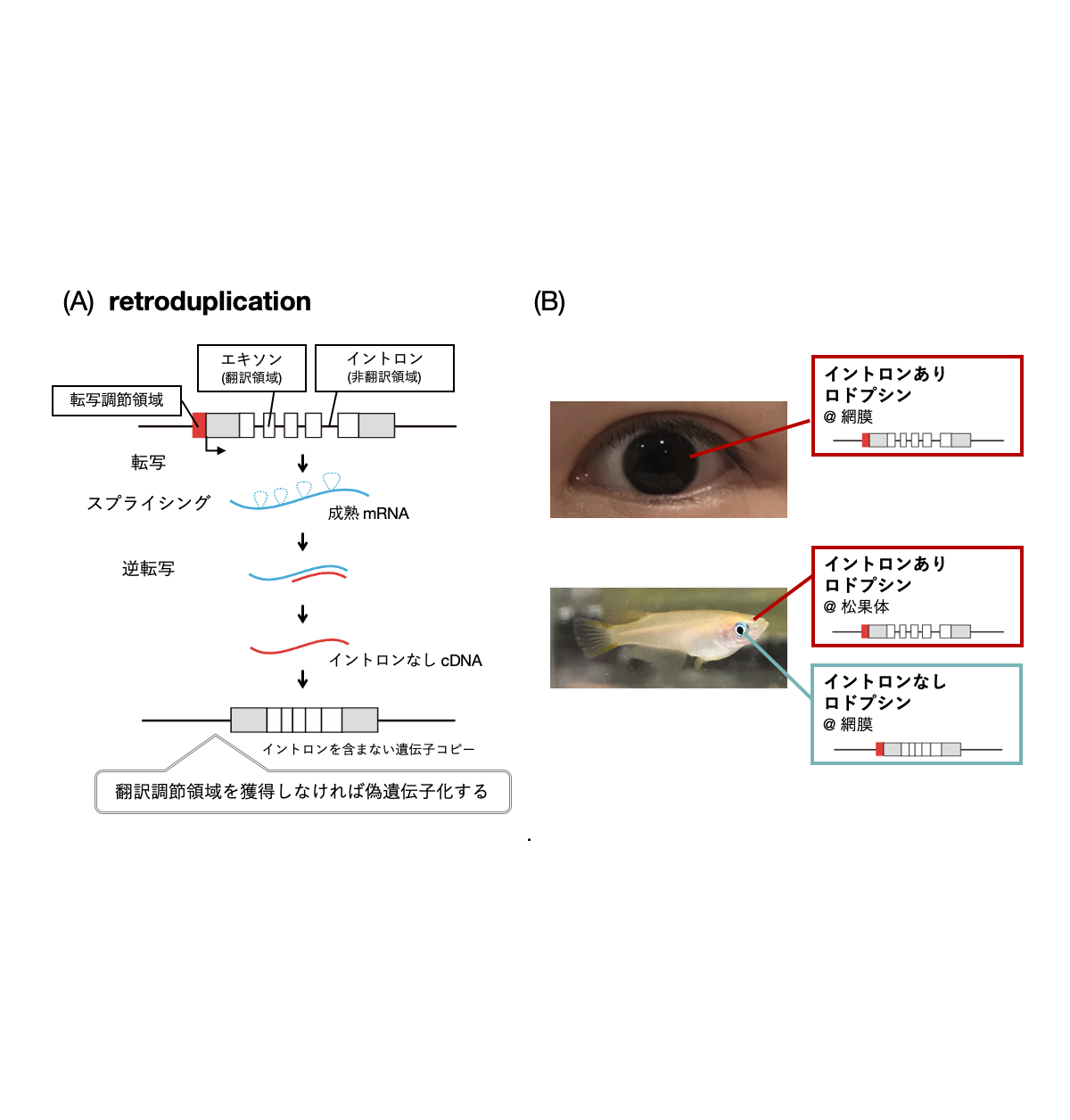
(A) retroduplicationによる遺伝子重複メカニズム
(B) 脊椎動物の多くはロドプシン遺伝子を一種類持ち、網膜で利用するが、真骨魚類は例外的に遺伝子構造の異なる複数種類のロドプシン遺伝子を持ち、イントロンを含まないものを網膜、イントロンを含むものを脳の松果体で利用する。
研究の背景
動物が外界環境を知覚する際に、光は重要な情報源であることが知られています。視覚によりものの形や色を認識することの重要性は言うまでもなく、それ以外にも光環境の変化から時刻や季節を認識することができます。このような多様な光受容に共通して機能する光センサータンパク質が、ロドプシンに代表されるオプシン類です。近年、動物のゲノム解析が進展してくると、動物によってオプシン遺伝子の数やレパートリーが大きく異なることがわかってきました。ヒトを含む哺乳類は10弱のオプシン遺伝子しか持っていませんが、鳥類・爬虫類・両生類は20遺伝子前後、魚類は30~40遺伝子を持っています。哺乳類でオプシン遺伝子の数が少ないのは、その祖先において夜行性であった時期があるためであると考えられています。これらのオプシン類は、アミノ酸配列に類似性があり、分子構造としても光受容のための発色団レチナールを結合するという共通性が見られます。このことから、様々な動物のもつオプシン遺伝子は元々1つの祖先型オプシン遺伝子に由来し、遺伝子重複と変異を繰り返すことによって現在の多様性が生まれたと考えられています。 オプシン類の遺伝子構造を確認すると、ほとんどのオプシン遺伝子は翻訳領域にイントロンを含みます。しかしごく少数ではありますが、イントロンを含まないオプシン遺伝子の存在も知られていました。このようなイントロンを含まない遺伝子は、retroduplicationという非常に珍しいユニークな遺伝子重複機構によって生じたと考えられています(図1(A))。retroduplicationでは祖先型のイントロンを含むオプシン遺伝子の成熟mRNAが鋳型として逆転写され、生じたcDNAがゲノムの別の領域へ挿入されることによって遺伝子重複が起こります。これは、ゲノム上の遺伝子を含むDNAのある領域のコピーが別のゲノム領域に直接挿入されるような一般的な遺伝子重複とは大きく異なります。しかし、retroduplicationにより生じた遺伝子コピーは通常近傍に発現調節領域を持たないため、機能的な遺伝子としては利用できない(偽遺伝子化)場合がほとんどであると考えられます。そのため、イントロンを含まないオプシン遺伝子が機能的に利用されている例は非常に珍しいと言えます。 本研究では、このようなイントロンを含まないオプシンとして、真骨魚類が持つロドプシンに着目しました(図1(B))。ロドプシンは通常、網膜の視細胞に発現し、視覚に関わる光センサーです。真骨魚類はロドプシン遺伝子を複数種類もち、イントロンを含まないものは網膜に、イントロンを含むものは網膜ではなく、時刻の認識に関わる光受容器官である脳の松果体に発現するということが知られていました。魚類以外の脊椎動物はイントロンを含むロドプシン遺伝子のみを持ち、網膜に発現させます。そのため、真骨魚類に至る進化的過程でイントロンを含まないロドプシン遺伝子が生じ、網膜で利用される一方、祖先型のイントロンを含む遺伝子は発現領域を網膜から松果体に変えたと考えられます。
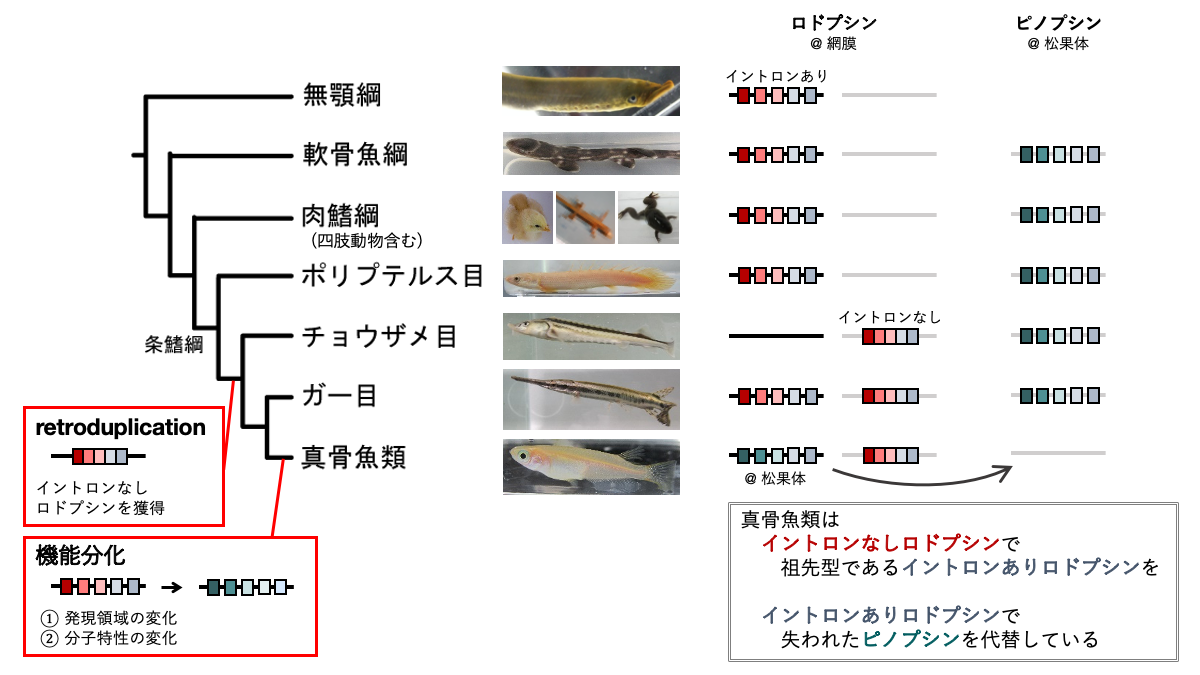
図2 条鰭綱におけるロドプシン遺伝子の段階的な獲得・分化モデル
ポリプテルス目は四肢動物と同様にイントロンを含むロドプシンを1種類持つ。ポリプテルス目の分岐後にretroduplicationによる遺伝子重複が起こり、イントロンなしロドプシンが獲得された。ガー目の分岐後、ピノプシンの喪失と並行して祖先型のイントロンありロドプシンの発現領域と分子特性が変化し、松果体特異的な光センサーへと分化した。
研究成果
そこで本研究では、ユニークな遺伝子重複メカニズムにより生じた真骨魚類のロドプシンが、どのような過程を経て眼と脳で多様な光受容機能に貢献するようになったのかを解明することを目指しました。そのために、条鰭綱において真骨魚類よりも古くに分岐したと考えられるポリプテルス・チョウザメ・ガーのロドプシン遺伝子を単離し、分子系統解析、網膜と脳での発現パターンの解析、タンパク質の性質の解析を行いました。これらの解析から、イントロンなしロドプシンはポリプテルス目の分岐後に生じて眼で働くようになり、一方祖先型のイントロンありロドプシンはガー目の分岐後に発現パターンやタンパク質の性質を変化させ、眼ではなく松果体で働くようになったと考えられました。また、イントロンありロドプシンが松果体で働くようになったことが、元々松果体で働いていた別のオプシンであるピノプシンを真骨魚類が失くすことにつながったのかもしれません。ピノプシンは、哺乳類を除く四肢動物や古代魚(ポリプテルス・チョウザメ・ガー)、軟骨魚類の松果体で働いています。ロドプシンとピノプシンはタンパク質の性質が比較的似ているため、真骨魚類はピノプシンの代わりにイントロンありロドプシンを松果体で使うようになった可能性があります。以上のことから、図2のような段階的な進化モデルが提唱できます。地球上に条鰭綱が登場する約4億年前から真骨魚類が登場する約2億年前の間に、ロドプシン遺伝子のユニークな遺伝子重複に端を発して、元々眼で働いていたロドプシン遺伝子が松果体で働くようになり、元々松果体で働いていたピノプシン遺伝子を失うというイベントが続けて起こったと考えられます。このように魚類は進化の過程において、通常の遺伝子重複だけではなくユニークで珍しい遺伝子重複も駆使してオプシン遺伝子を多様化させ、視覚だけに留まらない様々な機能に利用しています。このことは、水中における多彩な光環境に適応するために重要な意味を持ち、魚類の種の多様性の拡大にも貢献しているのかもしれません。