脳科学辞典(脳科学分野の約1,000個の用語を解説し、無償で公開しているサイト)でロドプシンについて解説しました。以下脳科学辞典「ロドプシン」より一部抜粋
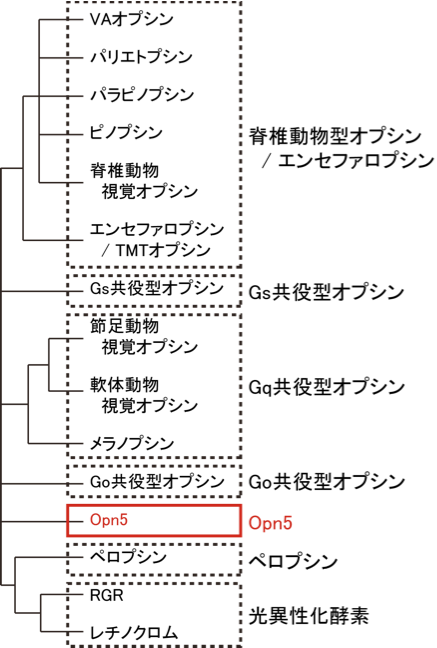
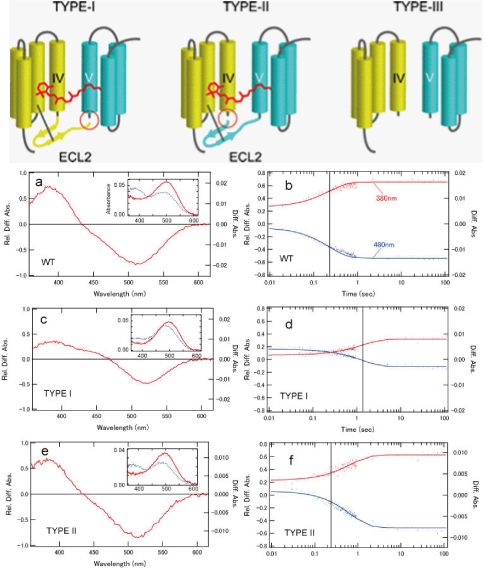
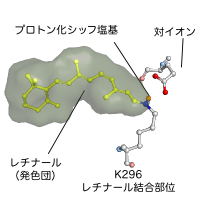
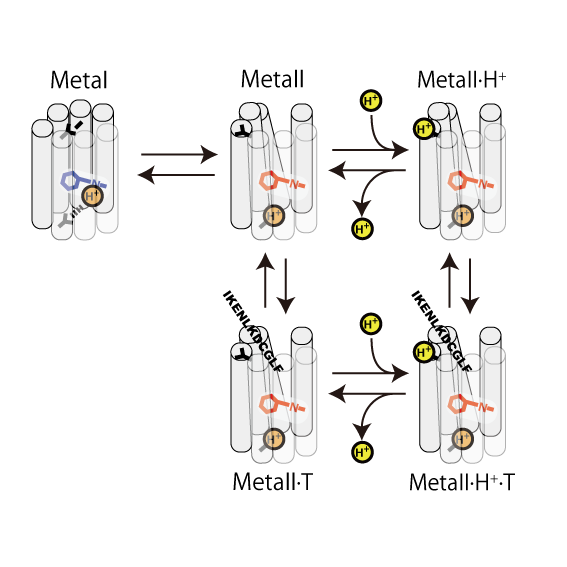
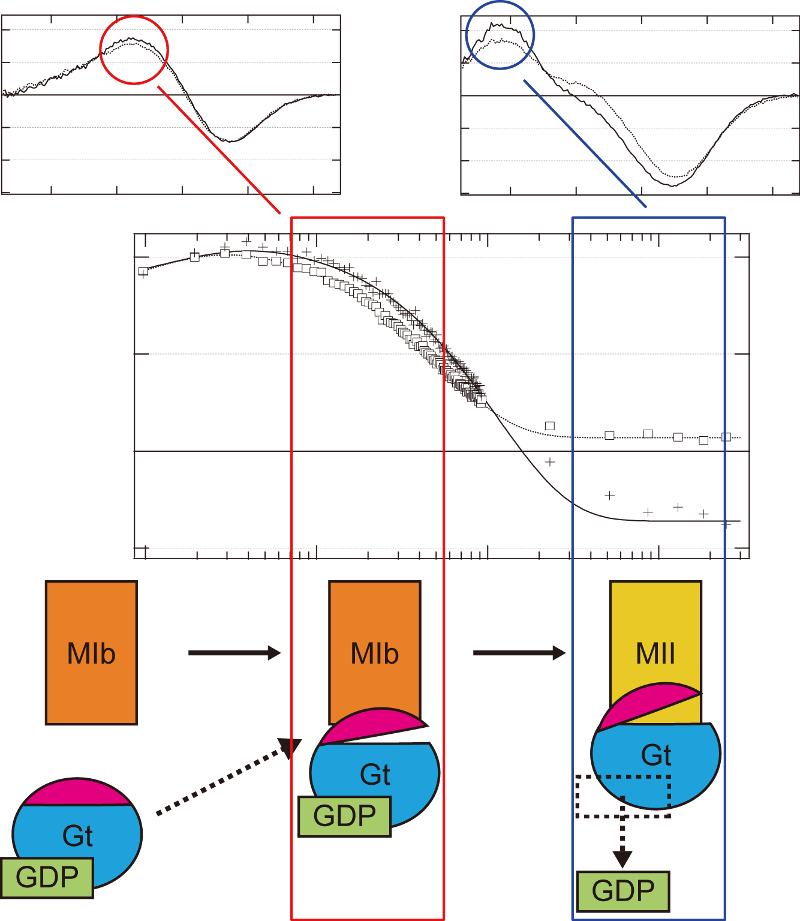
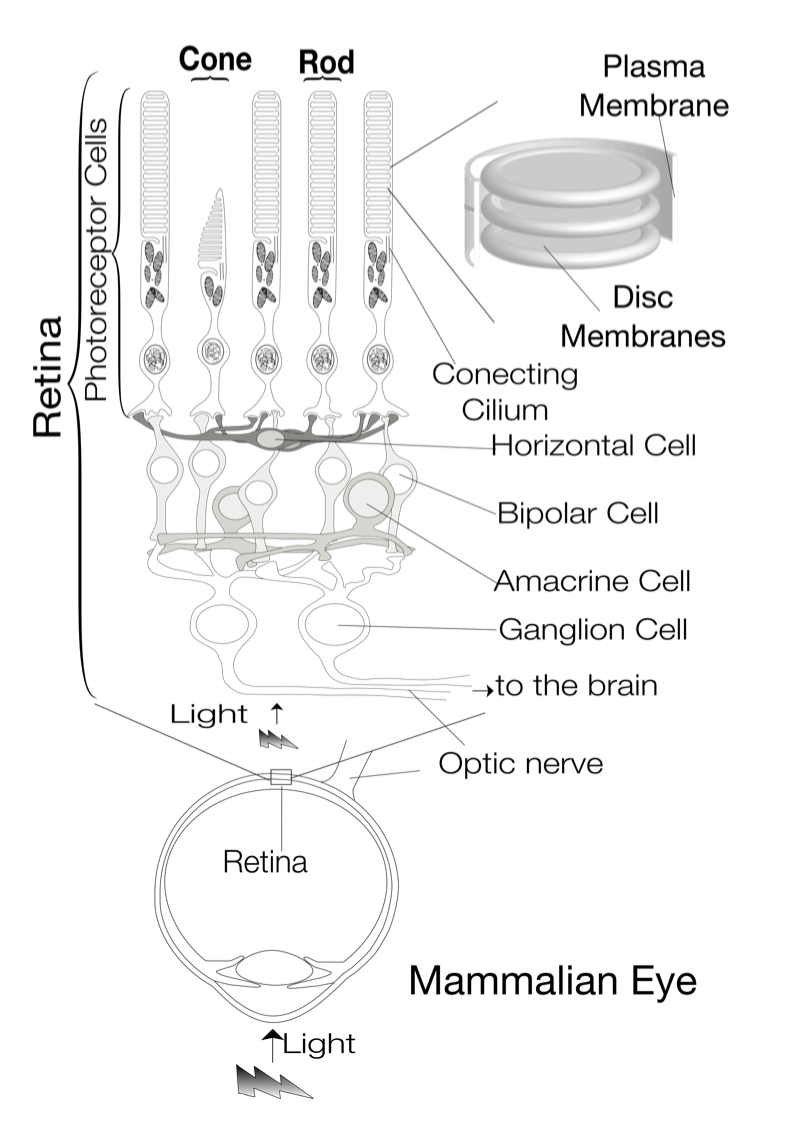
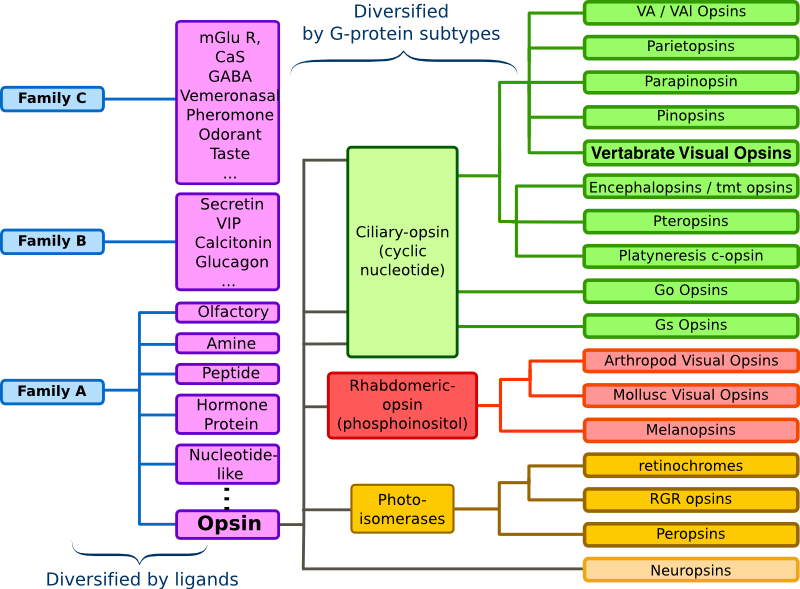
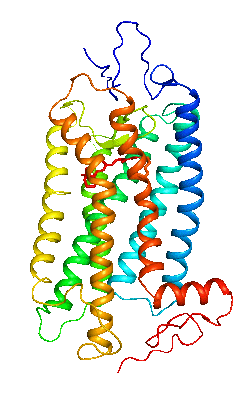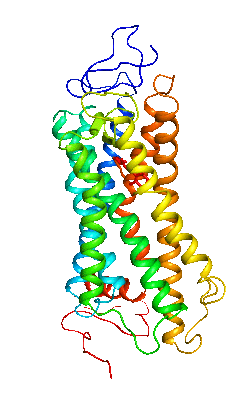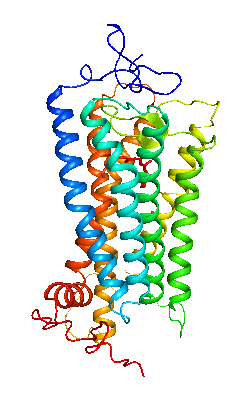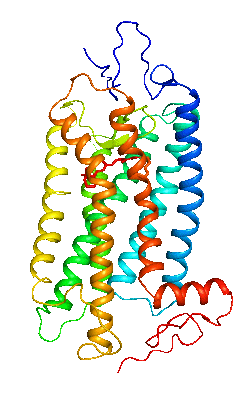
眼の網膜には光受容に特化した2種類の視細胞、桿体と錐体が含まれ、それぞれには光を受容するために特別に分化したタンパク質(光受容タンパク質)が含まれる。これらのタンパク質を視物質と呼び、特に桿体の視物質(桿体視物質)をロドプシンと呼ぶ。桿体には外節と呼ばれる光受容に特化した領域があり、その中には円盤膜(disk membrane)と呼ばれるパンケーキ状の膜構造がある。ロドプシンはこの膜構造の中に大量に埋め込まれて存在している。 ロドプシンはアポタンパク質と発色団レチナールより構成されており、レチナールが光を吸収することによって異性化しタンパク質部分の構造変化を起こし、Gタンパク質を介して細胞内シグナル伝達系を駆動する。光を吸収するという光受容体としての機能・特性がそのまま分子の物性を反映するプローブとして使えるため、ロドプシンは分光法によってその光反応過程が詳細に解析されており、Gタンパク質共役型受容体(GPCR)のなかでも最も研究が進んでいる受容体としても注目されている。また近年ではロドプシンに近縁なタンパク質(ロドプシン類あるいはオプシン類)が多く同定されており、さまざまな生物種の視覚の他に概日リズムの光同調などの視覚以外の生理機能を担っていることが明らかになっている。